

еуНДѓбЇЩњУќПЦбЇбЇдК аьБљ
еуНКМжн 310029
1 ЙњФкЭтбаОПЯжзД
ФтФЯНцЃЈArabidopsis thalianaЃЉЪЧвЛжжФЃЪНжВЮяЃЌОпгаЛљвђзщаЁЃЈ125 MbpЃЉЁЂЩњГЄжмЦкЖЬЕШЬиЕуЃЌЖјЧвЛљвђзщВтађвбОЭъГЩЃЈThe Arabidopsis Genomic Initiative, 2000ЃЉЁЃЭЌЪБЃЌФтФЯНцЪєЪЎзжЛЈПЦЃЈCruciferaeЃЉЃЌОпгаИпЕШжВЮяЕФвЛАуЬиЕуЃЌФтФЯНцбаОПжаЫљШЁЕУГЩЙћКмШнвзгУгкЦфЫќИпЕШжВЮяАќРЈХЉзїЮяЕФбаОПЃЌВњЩњжиДѓЕФОМУаЇвцЃЌЬиБ№ЪЧЪЎзжЛЈПЦжаЛЙгааэЖрживЊЕФОМУзїЮяЃЌгыШЫРрЕФЩњВњЩњЛюУмЧаЯрЙиЃЌвђДЫФПЧАФтФЯНцЕФбаОПдНРДдНЖрЕиЪмЕНЙњМЪжВЮябЇМАИїЙњеўИЎЕФжиЪгЁЃ
ДгвХДЋбЇЕФЙлЕуРДПДЃЌЛљвђПЫТЁЕФЭООЖПЩИХРЈЮЊе§ЯђвХДЋбЇКЭЗДЯђвХДЋбЇСНжжЁЃе§ЯђвХДЋбЇЭООЖжИЕФЪЧЭЈЙ§БЛПЫТЁЛљвђЕФВњЮяЛђБэЯжаЭЭЛБфШЅНјааЃЛЗДЯђвХДЋбЇЭООЖдђжИЕФЪЧвРОнБЛПЫТЁЛљвђдкШОЩЋЬхЩЯЕФЮЛжУРДЪЕЯжЁЃЫфШЛвЛаЉФЃЪНЩњЮяЃЈШчФтФЯНцЃЉЕФЛљвђзщВтађвбОЭъГЩЃЌЕЋЛЙга40%ЕФЛљвђЃЈдкФтФЯНцжаЃЉЕФЙІФмЛЙЪЧЮДжЊЕФЁЃ
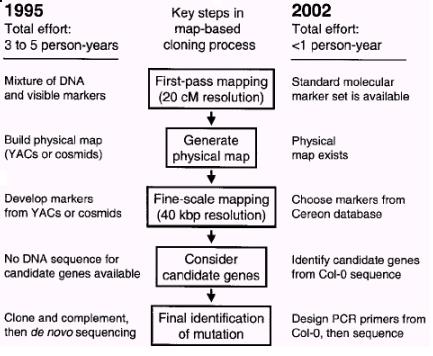
ЭМ1 ЭМЮЛПЫТЁЫљашХЌСІЕФБШНЯЃЈ1995ФъКЭ2002ФъЃЉЃЈJanderЕШЃЌ 2002ЃЉ
ЭМЮЛПЫТЁЃЈmap-based cloningЃЉгжГЦЖЈЮЛПЫТЁЃЈpositional cloningЃЉЃЌ1986ФъЪзЯШгЩНЃЧХДѓбЇЕФAlan CoulsonЬсГіЃЈCoulsonЕШЃЌ1986ЃЉЃЌгУИУЗНЗЈЗжРыЛљвђЪЧИљОнФПЕФЛљвђдкШОЩЋЬхЩЯЕФЮЛжУНјааЕФЃЌЮоашдЄЯШжЊЕРЛљвђЕФDNAађСаЃЌвВЮоашдЄЯШжЊЕРЦфБэДяВњЮяЕФгаЙиаХЯЂЁЃЫќЪЧЭЈЙ§ЗжЮіЭЛБфЮЛЕугывбжЊЗжзгБъМЧЕФСЌЫјЙиЯЕРДШЗЖЈЭЛБфБэаЭЕФвХДЋЛљДЁЁЃНќМИФъРДЫцзХФтФЯНцЛљвђзщВтађЙЄзїЕФЭъГЩЃЌИїжжЗжзгБъМЧЕФШеЧїЗсИЛКЭИїжжЪ§ОнПтЕФЭъЩЦЃЌдкФтФЯНцжаПЫТЁвЛИіЛљвђЫљашвЊЕФХЌСІвбОДѓДѓМѕЩйСЫЃЈЭМ1ЃЉЁЃ
ФПЧАЭъГЩећИіФтФЯНцЕФЭМЮЛПЫТЁЙ§ГЬДѓдМашвЊвЛФъЪБМфЁЃдкетИіЙ§ГЬжаЃЌЮвУЧДгЩИбЁЭЛБфЬхПЊЪМЃЌж№НЅевЕНКЭБэаЭЯрЙиЕФЛљвђЁЃетКЭЗДЯђвХДЋбЇЕФЗНЗЈе§КУЯрЗДЁЃЭМЮЛПЫТЁФмЪЕЯжЃЌЙиМќдкгкШЋЛљвђзщВтађМЦЛЎЕФЭъГЩКЭИїжжЗжзгБъМЧЕФЗЂЯжЁЃетаЉЪ§ОнБЛДЂДцдкзЈУХЕФЪ§ОнПтжаЃЈБэ1ЃЉЃЈLukowitzЕШЃЌ 2000ЃЉЁЃдкФтФЯНцжаЕФЭМЮЛПЫТЁЃЌдкКмДѓГЬЖШЩЯЕУвцгкЖдCol-0ЩњЬЌаЭВтађЕФЭъГЩЃЌвђЮЊЫќЪЧдкбаОПФтФЯНцЪБзюГЃгУЕФЩњЬЌаЭЁЃ
ЪЕЯжЛљвђЭМЮЛПЫТЁЕФЙиМќЪЧЩИбЁгыФПБъЛљвђСЌЫјЕФЗжзгБъМЧЁЃЪЕжЪЩЯЃЌЗжзгБъМЧЪЧвЛИіЬивьЕФDNAЦЌЖЮЛђФмЙЛМьГіЕФЕШЮЛЛљвђЃЌЖдЦфгааЇЕиРћгУМДПЩДяЕНЭМЮЛПЫТЁЛљвђжЎФПЕФЁЃЦљНёЮЊжЙЃЌвбгаМИЪЎжжММЪѕПЩгУгкЗжзгБъМЧЕФЩИбЁЃЈWangЕШЃЌ2000ЃЉЁЃЦфжазюЮЊГЃгУЕФЪЧМђЕЅађСаГЄЖШЖрЬЌадЃЈSSLPsЃЉЃЈLukowitzЕШЃЌ 2000ЃЛ ChoeЕШЃЌ 2002ЃЛ Gonzalez-GuzmanЕШЃЌ 2002ЃЉЁЃКЭЕЅКЫмеЫсЖрЬЌадЃЈSNPsЃЉЃЈRafalskiЃЌ 2002ЃЉЁЃSSLPЪЧЛљгкPCRЕФЗжзгБъМЧЃЌдкФтФЯНцЛљвђзщжагаНЯЖрЗжВМЃЌЖјЧвЪЧЙВЯдадЕФЃЌЫќЕФМьВтЗЧГЃжБНгЃЌЕЋЪЧЮвУЧашвЊЩшМЦв§ЮяРДМьВтМйЖЈЕФSSLPБъМЧЃЛЖдSNPsБъМЧЕФМьВтвВБШНЯжБНгЃЌЫќЪЧФтФЯНцВЛЭЌЩњЬЌаЭжЎМфЛљвђзщжаЕФЕЅИіКЫмеЫсЕФВюБ№ЃЌетаЉВюБ№ЕФКЫмеЫсЭЈГЃЮЛгкВЛБрТыЧјгђЃЈPetersЕШЃЌ 2003ЃЉЁЃзюГЃМћЕФгУгкМьВтSNPsБъМЧЕФЗНЗЈжївЊЪЧМєЧаРЉдіЖрЬЌадађСаЃЈCAPSЃЉЃЌЫќвВЪЧЛљгкPCRЕФЁЃСэЭтЃЌвЛжжИќЮЊгааЇЕФЗНЗЈбмЩњЕФCAPSЃЈdCAPSЃЉЃЈNamЕШЃЌ 1989ЃЛ Michaels КЭ AmasinoЃЌ 1998ЃЉПЩАбШЮКЮвбжЊЕФЕуЭЛБфзїЮЊЗжзгБъМЧЃЌжЛвЊдкPCRЪЧв§ШыВЛХфЖдЕФв§ЮяЃЌЪЙРЉдіЕФађСадквЛИіЩњЬЌаЭжаОпгаЯожЦадУИЧаЮЛЕуЃЌЖјдкСэвЛЩњЬЌаЭжаУЛгаЃЌвдаЮГЩЖрЬЌадЁЃ
ЭМЮЛПЫТЁЗЈЫцзХЯрЙиХфЬзММЪѕЃЈађСаЪ§ОнПтЁЂЗжзгБъМЧЕШЃЉЕФШеНЅГЩЪьЃЌаэЖрФтФЯНцМАвЛаЉХЉзїЮяЕФЛљвђвбБЛГЩЙІЕФПЫТЁЃЈБэ2ЃЉЁЃБОЮФФтЖдЭМЮЛПЫТЁЕФбаОПНјеЙзівЛНщЩмЃЌвдЦкЖджВЮявХДЋг§жжКЭЗжзгЩњЮябЇбаОПгаЫљАяжњ
Бэ1 ФтФЯНцЭјТчзЪдД
|
ЭјеО
|
ЭјжЗ
|
|
Supplemental material for this paper
|
http://carnegiedpb.stanford.edu/methods/ppsuppl.html
|
|
Nottingham Stock CentreЃЈU.K.ЃЉ
|
http://nasc.nott.ac.uk/
|
|
Recombinant Inbred map
|
http://nasc.nott.ac.uk/new_ri_map.html
|
|
Ohio Stock CenterЃЈU.S.A.ЃЉ
|
http://aims.cps.msu.edu/aims/
|
|
TAIR database*ЃЌ homepage
|
http://www.arabidopsis.org
|
|
Recombinant Inbred mapЃЈmirror siteЃЉ
|
http://www.arabidopsis.org/cgi-bin/maps/Riintromap
|
|
CAPS markers
|
http://www.arabidopsis.org/aboutcaps.html
|
|
Sequence table
|
http://www.arabidopsis.org/cgi-bin/maps/Seqtable.pl
|
|
SNP collection
|
http://www.arabidopsis.org/SNPs.html
|
|
CEREON collection of polymorphisms
|
http://www.arabidopsis.org/cereon
|
|
SSLP markers
|
http://genome.bio.upenn.edu/SSLP_info/SSLP.html
|
|
TIGRЃЌ genome annotations
|
http://www.tigr.org/tdb/athl/htmls/index.html
|
|
Database of Ler sequences
|
http://www.tigr.org/tdb/atgenome/Ler.html
|
|
Kasuza DNA Research InstituteЃЌ genome annotations
|
http://www.kazusa.or.jp/kaos/
|
|
MIPS genome annotations
|
http://websvr.mips.biochem.mpg.de/proj/thal/
|
|
SINS database of transposon insertions
|
http://www.jic.bbsrc.ac.uk/sainsbury-lab/jonathan-jones/jjhome.htm
|
|
*зЂЃКThe Arabidopsis Information Resource ЃЈTAIRЃЉ
|
|
2 ЭМЮЛПЫТЁЕФвЛАуЙ§ГЬ
вђЮЊгаСЫФтФЯНцЕФЛљвђзщађСаКЭИпУмЖШЕФвХДЋБъМЧЃЌЭМЮЛПЫТЁЙ§ГЬОЭБфЕУЯрЖджБНгЁЃЭМ2Р§ОйСЫвЛжжИпаЇЕФФтФЯНцЭМЮЛПЫТЁЗНЗЈЁЃДгЛљгкCol-0КЭLerвХДЋБГОАЕФЭЛБфЬхГіЗЂЃЌЮвУЧгаПЩФмдкДѓдМвЛФъЪБМфФкевГігыетИіЭЛБфЯрЙиЕФЛљвђЃЌетЦфжажївЊКФЪБМфЕФЪЧЮхИіжВЮяЃЈФтФЯНцЃЉЕФЩњГЄжмЦкЃЈЮвУЧМйЖЈУПИіжмЦкЮЊСНИідТЃЉЁЃ
зїЮЊзїЭМЙ§ГЬЕФЕквЛВНЃЌЭЛБфЬхжВжъНЋКЭСэЭтвЛИіЩњЬЌаЭЃЈCol-0ЛђепLerЃЉЕФжВжъдгНЛЁЃдкДѓЖрЪ§ЧщПіЯТЃЌгУгкдгНЛЕФЭЛБфЬхжВжъЪЧзїЮЊИИБОЛЙЪЧФИБОЪЧУЛгаЙиЯЕЕФЁЃШЛКѓВЅжжF1ДњжжзгЁЃдкF1ДњжВЮяЕФЩњГЄЙ§ГЬжаЃЌЮвУЧОЭгаПЩФмРДЖдЦфБэЯжаЭКЭЛљвђаЭНјааЗжЮіЁЃF1ДњжВЮяЕФБэаЭЕФГіЯжЛђепЯћЪЇНЋЯдЪОзХЮвУЧЫљбаОПЕФЭЛБфЪЧЯдадЕФЛЙЪЧвўадЕФЁЃзюКУЭЈЙ§ЖдвЛаЉБъМЧЕФЗжЮіРДШЗШЯF1ДњжВЮяЪЧдгКЯЬхЃЌЖјЧвдкдгНЛЙ§ГЬжаЮвУЧУЛгаЗИДэЮѓЁЃЕБШЛвВгаБивЊШЗШЯдРДЕФЩњЬЌаЭБГОАЁЃ
Бэ2 гУЭМЮЛПЫТЁЗНЗЈЕУЕНЕФФтФЯНцМАвЛаЉХЉзїЮяЕФЛљвђ
|
Лљвђ
|
ЭЛБфБэаЭ
|
ЛљвђЭЌдДађСа
|
AB13 |
ЭбТфЫсВЛУєИа
|
гёУззЊТМзг
|
|
FID3
|
НЕЕЭбЧгЭЫсБЅКЭЖШ
|
ЯИОњШЅБЅКЭЫсУИ
|
|
AXR1
|
ЩњГЄЫиПЙад
|
ЗКЫиN ЖЫЛюадУИ
|
|
ETR1
|
ввЯЉПЙад
|
ЫЋвђзгЕїНкзг
|
|
ABI1
|
ЭбТфЫсВЛУєИа
|
ИЦЕїЕААзСзЫсЛЏУИ
|
|
DET1
|
ЛЦЛЏЫ№ЩЫЗДгІ
|
аТКЫЕААз
|
|
RPS2
|
ПЙВЁ
|
аТаЭИЛКЌССАБЫсЕФЕААзУИ
|
|
RPM1
|
ПЙВЁ
|
МЄУИ
|
|
RSW1
|
ЯЫЮЌЫиКЯГЩУИ
|
ЯИАћЩЋЫиP450 МвЯЕ
|
|
ZLL
|
ЕїНкжаОЅЗжЩњ
|
ЯИАћХпЬЅЗЂг§ЕААз
|
|
PRT1
|
вжжЦАћМфЕААзНЕНт
|
ПижЦжВЮяN ЖЫДњаЛ
|
|
Tornadol
|
жВжъЖЬЛЏ
|
——
|
|
IFL1
|
е§ГЃЕФЮЌЙмЪјМфЯЫЮЌЗжЛЏЪмзш
|
ССАБЫсРСДЕААз
|
|
ARA1
|
ЪїНКШЉЬЧМЄУИЛюадЩЅЪЇ
|
АыШщЬЧМЄУИЛљвђМвзх
|
|
VTC2
|
ЮЌЩњЫиCКЯГЩВЛзу
|
ЙћгЌЕААзCG3552ЃЌЯпГцЕААзC10F3.4ЃЈЙІФмЮДжЊЃЉ
|
|
AST
|
жжЦЄЛЈЧрмеАпЕу
|
ЛЈЧрмеЩњЮяКЯГЩЭООЖжаЕФЖўЧтЛЦЭЊДМ-4-ЛЙдУИ
|
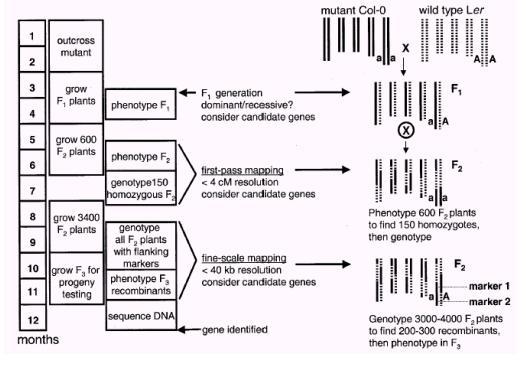
ЭМ2 ЭМЮЛПЫТЁЙ§ГЬЪОвтЭМЃЈJanderЕШЃЌ 2002ЃЉ
F1ДњжВЮяздНЛЕУЕНF2ДњжжзгЃЌДѓдМВЅжж600ИіИіЬхвдНјааЭЛБфЛљвђЕФДжЖЈЮЛЃЈfirst-pass mappingЃЌЭМ2ЃЉЁЃдкЦфЩњГЄЙ§ГЬжаЃЌЮвУЧПЩШЗЖЈЦфБэаЭЃЌДѓдМга150ИіИіЬхБЛШЯЮЊЪЧДПКЯЬхЃЈдквўадЭЛБфЕФЧщПіЯТЪЧДПКЯЭЛБфЬхЃЌдкЯдадЭЛБфЕФЧщПіЯТЪЧДПКЯвАЩњаЭЃЉЁЃШЛКѓДгет150ИіИіЬхЕФвЖзгЛђепЦфЫќзщжЏжажЦБИDNAгУгкЛљвђаЭЗжЮіЁЃЦ№ЯШгУЗжВМгкФтФЯНцЮхЬѕШОЩЋЬхЩЯЕФ25ИіБъМЧЃЈЯрСкЕФСНИіБъМЧжЎМфДѓдМЯрОр20 cMЃЉНјааЗжЮіЃЌШЗЖЈЭЛБфЛљвђЪЧКЭФФИіЛђепФФМИИіБъМЧЪЧСЌЫјЕФЃЌШЛКѓгУШ§ЕуВтНЛЕФЗНЗЈРДЖЈвхвЛИіАќКЌЭЛБфЛљвђЕФДѓдМ20 cMЕФвХДЋМфИєЁЃвЛЕЉетбљЕФвЛИівХДЋМфИєБЛЖЈвхжЎКѓЃЌНгЯТРДЕФЙЄзїОЭЪЧв§ШыаТЕФБъМЧАбетИіМфИєЫѕаЁЕНДѓдМ4 cMЁЃвЛАуРДЫЕЃЌРћгУ150ИіF2ДњИіЬхЪЧдкКмДѓГЬЖШЩЯФмевЕНетбљвЛИівХДЋМфИєЕФЃЌОрРыЭЛБфЛљвђзюНќЕФСНИіЗжзгБъМЧНЋзїЮЊВрУцБъМЧЖјгУгкЯТУцЕФНјвЛВНЗжЮіЁЃ
ЯТвЛВНЮвУЧНЋВЅжжвЛИіИќДѓЕФF2ДњШКЬхгУгкЭЛБфЛљвђЕФОЋЯИЖЈЮЛЃЈfine-resolution mappingЃЌЭМ2ЃЉЁЃзюжеФПБъЪЧНЋАќКЌЭЛБфЛљвђЕФвХДЋМфИєЫѕаЁЕН40 KbЩѕжСИќаЁЃЈетдкФтФЯНцжаДѓдМЪЧ0.16 cMЃЉЁЃЯдШЛгУгкзїЭМЕФF2ДњжВЮядНЖрЃЌОЭдНФмОЋШЗЕиЖЈЮЛЭЛБфЛљвђЁЃвЛАуашвЊ3000~4000ИіF2ДњжВЮяИіЬхЃЈАќРЈДжЖЈЮЛЪБЕФ600ИіF2ДњжВЮяИіЬхЃЉРДОЋШЗЕиЖЈЮЛЭЛБфЛљвђЁЃЕЋЪЧвВгаКмЖрЭМЮЛПЫТЁЙ§ГЬгУСЫЩйгк3000ИіF2ДњжВЮяИіЬхОЭГЩЙІЕиЖЈЮЛСЫЭЛБфЛљвђЃЈLukowitzЕШЃЌ 2000ЃЉЁЃЕЋЪЧетЭљЭљвЊУАвђЮЊзїЭМШКЬхВЛЙЛДѓдйвЛДЮжжжВF2ДњжВЮяЖјбгГЄећИізїЭМЙ§ГЬЕФЪБМфЕФЗчЯеЁЃ
дкетИіДѓдМ4 cMЕФвХДЋМфИєФкевЕНгыЭЛБфИќНєУмСЌЫјЕФЗжзгБъМЧЃЌвЛАуЧщПіЯТФмдкЭЛБфСНВревЕНЯрОраЁгк40 KbЕФСНИіЗжзгБъМЧЁЃвЛЕЉетбљЕФСНИіЗжзгБъМЧБЛевЕНжЎКѓЃЌОЭПЩвдЭЈЙ§ВтађРДевЕНЭЛБфЛљвђЁЃвЛжжгааЇЕФЗНЗЈЪЧЩшМЦPCRв§ЮяРДРЉдіИВИЧет40 KbЕФЖрИіжиЕўЕФ500 bpЕФЦЌЖЮЁЃНЋетаЉЦЌЖЮВтађКѓЦДНгЦ№РДвдЕУЕНећИі40 KbЕФађСаЃЌШЛКѓНЋЫќгывАЩњаЭжВЮяЃЈCol-0ЛђепLerЃЉЕФађСаНјааБШЖдЃЌетОЭПЩвдевЕНетИіЧјгђжаЕФЖрИіЛљвђЁЃДгвЛЯЕСаКюбЁЛљвђжаМјЖЈЛљвђЪЧЖЈЮЛПЫТЁММЪѕЕФзюКѓвЛИіЙиМќЛЗНкЁЃЯждкзюГЃгУЕФЗНЗЈЪЧгУКЌгаФПБъЛљвђЕФДѓЦЌЖЮПЫТЁШчBACПЫТЁЛђYACПЫТЁШЅЩИбЁcDNAЮФПтЃЌВЂВщбЏЩњЮяЪ§ОнаХЯЂПтЃЌД§евГіКюбЁЛљвђКѓЃЌАбетаЉКюбЁЛљвђНјааЯТСаЗжЮівдШЗЖЈФПБъЛљвђЃКЃЈ1ЃЉОЋШЗЖЈЮЛЗЈМьВщcDNAЪЧЗёгыФПБъЛљвђЙВЗжРыЃЛЃЈ2ЃЉМьВщcDNAЪБПеБэДяЬиЕуЪЧЗёгыБэаЭвЛжТЃЛЃЈ3ЃЉВтЖЈcDNAађСаЃЌВщбЏЪ§ОнПтЃЌвдСЫНтИУЛљвђЕФЙІФмЃЛЃЈ4ЃЉЩИбЁЭЛБфЬхЮФПтЃЌевГіDNAађСаЩЯЕФБфЛЏМАгыЙІФмЕФЙиЯЕЃЛЃЈ5ЃЉНјааЙІФмЛЅВЙЪЕбщЃЌЭЈЙ§зЊЛЏЭЛБфЬхЙлВьЭЛБфЬхБэаЭЪЧЗёЛжИДе§ГЃЛђЗЂЩњдЄЦкЕФБэаЭБфЛЏЁЃЙІФмЛЅВЙЪЕбщЪЧзюжБНгЁЂзюжеМјЖЈЛљвђЕФЗНЗЈЁЃРћгУаТаЫЕФRNAИЩШХЃЈRNAiЃЉвВПЩгааЇЕиШЗЖЈФПЕФЛљвђЁЃ
3 ДцдкЕФЮЪЬт
ЭМЮЛПЫТЁвВгаЦфздЩэЕФОжЯоадЃЌдкФГаЉЧщПіЯТЃЌОЭКмФбЛђепВЛФмЭЈЙ§ЭМЮЛПЫТЁММЪѕРДЖЈЮЛЛљвђЁЃ
дкЗжЮіздШЛЗЂЩњЕФБфвьЕФЪБКђЃЌЮвУЧзюгаПЩФмгіЕНЕФИДдгЧщПіЪЧвЛИіИјЖЈЕФадзДЪЧгЩВЛжЙвЛИіЕФЛљвђЮЛЕуПижЦЕФЁЃР§ШчЃЌдкKashmir-1ЃЈгаПЙадЕФЃЉКЭColumbiaЃЈУєИаЕФЃЉжъЯЕжЎМфЕФдгНЛЪЕбщжаЃЌЮвУЧЗЂЯжЗлзДУЙОњПЙадЛљвђжСЩйЩцМАШ§ИівХДЋЮЛЕуЃЌЫќУЧЪЧвдИНМгЕФЗНЪНЦ№зїгУЕФЃЈI.WilsonЃЌ C. SchiffЃЌ КЭ S. SomervilleЃЌИіШЫНЛСїЃЉЁЃЖдетаЉПЙадЛљвђжаЕФШЮКЮвЛИізїОЋЯИЖЈЮЛЖМвЊЧѓНЕЕЭзїЭМШКЬхЕФвХДЋИДдгадЃЌР§ШчЭЈЙ§ДДдьжЛгавЛИіЮЛЕуБЃГжЖрЬЌадЕФжизщНќНЛЯЕЁЃдкФтФЯНцЕФжъЯЕжЎМфдгНЛЪБЃЌКмЖржжадзДЪЧгЩвЛИіЛђЖрИівХДЋЮЛЕуПижЦЕФЃЌЦфжаАќРЈПЊЛЈЪБМфЃЌжжзгДѓаЁЃЌЖЌУпЃЌЩњРэНкТЩЃЌДЮЩњДњаЛвдМАБэЦЄУЋЕФУмЖШЃЈзлЪіМћAlonso-Blanco КЭ KoornneefЃЌ2000ЃЉЁЃЮоТлКЮЪБЃЌЕБгАЯьетаЉадзДЕФздШЛЛђепгеЕМЕФЭЛБфБЛЖЈЮЛЕФЪБКђЃЌЕкЖўЮЛЕуаоЪЮГЩЗжЛсИЩШХетаЉЗжЮіЁЃ
БэЙлЃЈЩЯЮЛЃЉвХДЋЭЛБфетИіЪѕгяЪЧУшЪівЛИіЛљвђдкБэДяКЭЙІФмЩЯЕФПЩвХДЋИФБфЃЌЖјВЛЩцМАDNAађСаЕФИФБфЃЈзлЪіМћWolffe КЭ Matzke,1999ЃЉЃЌетЪЧЭМЮЛПЫТЁЙЄГЬжагжвЛИіПЩФмЕФИДдгЧщПіЁЃвбгаЮФЯзКмКУЕижЄУїЕФЪЧЛЈЗЂг§ЛљвђSUPERMANЕФКѓЩњclark kantЕШЮЛЛљвђЃЈJacobson КЭ MeyerowitzЃЌ1997ЃЉЁЃетаЉЕШЮЛЛљвђЪЧПЩвХДЋЕФЃЌЕЋЫќУЧВЛЮШЖЈгавЛИіаЁЕФЛиИДТЪЁЃЫќУЧдкSUPERMANЛљвђЕФDNAађСажаЖМОпгаЯрЫЦЕФАћрзрЄМзЛљЛЏЯжЯѓЃЌНсЙћЃЌгаПЩФмМѕЩйСЫSUPERMANЛљвђзЊТМзгЕФБэДяЁЃЫќУЧжаУЛгавЛИіЪЧКЭSUPERMANЕФDNAађСаИФБфСЊЯЕдквЛЦ№ЕФЃЛОЁЙмШчДЫЃЌЫќУЧФмБЛДјгаSUPERMANЛљвђЕФзЊЛљвђЫљВЙГфЁЃФПЧАЃЌЖдгкетжжБэЙлвХДЋЭЛБфЪЧдѕУДВњЩњЕФвдМАЫќУЧГіЯжЕФЦЕТЪжЊЕРЕФВЛЖрЁЃ
ЙигкШОЩЋЬхЩЯЮЛЕуЕФЮяРэКЭвХДЋОрРыЕФБШжЕЪЧБфЛЏЕФЁЃЭЈГЃетжжБфЛЏЪЧБШНЯаЁЕФЃЌЖдзїЭМЕФЗжБцТЪвВжЛгаНЯаЁЕФгАЯьЃЈCopenhaverЕШЃЌ1998ЃЉЁЃЕЋЪЧЃЌгажЄОнБэУїгааЉШОЩЋЬхЧјгђЪЧР§ЭтЕФЁЃР§ШчЃЌЖдGURKEЛљвђЕФЭМЮЛПЫТЁОЭЗЧГЃРЇФбЃЌетИіЛљвђЕФЖЈЮЛНгНќгкЕквЛЬѕШОЩЋЬхЕФзХЫПСЃЃЛдкзХЫПСЃИННќжизщЪЧбЯИёЯожЦЕФЃЌЪЙЕУЖдЫќОЋЯИЖЈЮЛЕФХЌСІЗЧГЃЮоаЇЁЃЖјЧвЃЌдкетИіЧјгђжажиИДDNAЕЅдЊЕФЙуЗКЗжВМЪЙЮвУЧБцШЯГіЩЂВМЕФЕЅПНБДађСаЃЌетаЉЕЅПНБДађСаФмВњЩњгавЩЮЪЕФвХДЋБъМЧЃЈR. Torres RuizЃЌИіШЫНЛСїЃЉЁЃетИіЗЂЯжЪЧОЙ§ЖдЕкЖўЬѕШОЩЋЬхЩЯЕФЮяРэКЭвХДЋОрРыжЎМфЕФБШжЕЕФЯЕЭГЕиЗжЮіжЎКѓШЗШЯЕФЃЈLinЕШЃЌ1999ЃЉЁЃЖдетЬѕШОЩЋЬхЕФМИКѕШЋађСаЃЌ1%жизщЕФвХДЋОрРыЯрЕБгк100~400 KbЕФЮяРэОрРыЃЌЦНОљЪЧ250 KbЁЃШЛЖјзХЫПСЃЧјгђЪЧвЛИіЯджјЕФР§ЭтЃЌдкетРя1%жизщЕФвХДЋОрРыЯрЕБгк1000~2500 KbЁЃПДРДжЕЕУжИГіЕФЪЧдкЯжДцЕФЮяРэЭМЦзжаЃЌФтФЯНцЕФЮхИізХЫПСЃЪЧУЛгавЛИіБЛЭъШЋИВИЧЕФЁЃзюНќЖдзХЫПСЃЧјгђЕФЗжЮіЯдЪОетаЉЧјгђЭЈГЃАќКЌжиИДЕФDNAКЭМИКѕВЛКЌБэДяЕФЛљвђЃЈCopenhaverЕШЃЌ1999ЃЉЁЃвђДЫЃЌгЩгкНгНќзХЫПСЃЃЌгІИУУЛгаФтФЯНцЛљвђЪЧВЛЗўДгЭМЮЛПЫТЁВпТдЕФЁЃ
Г§СЫзХЫПСЃЃЌЕкЖўЬѕШОЩЋЬхЩЯвВгавЛИіаЁЦЌЖЮЩЯ1%жизщЕФвХДЋОрРыЯрЕБгк1000 KbЩѕжСИќЖрЁЃИљОнЭЦВтЃЌЙлВьЕНЕФЕЭжизщТЪЯжЯѓПЩФмЪЧгЩгкБЛгУгкзїЭМЗжЮіЕФжъЯЕЕФDNAађСаЕФжиХХЃЈLinЕШЃЌ1999ЃЛMayerЕШЃЌ1999ЃЉЁЃЕкЖўКЭЕкЫФЬѕШОЩЋЬхЕФDNAађСаЕФБШНЯЯдЪОгааЉЛљвђЦЌЖЮЪЧдкетСНЬѕШОЩЋЬхжЎМфБЛИДжЦЕФЃЈЦфжавЛИіЦЌЖЮЕФДѓаЁЪЧ4.6 MbЃЉЃЌЛЙгавЛИіДгЯпСЃЬхЛљвђзщЯђЕкЖўЬѕШОЩЋЬхзЊвЦЕФDNAЦЌЖЮЃЈLinЕШЃЌ1999ЃЉЁЃетаЉЗЂЯжЧхГўЕижЄУїСЫФтФЯНцЛљвђзщЕФНсЙЙЪЧПЩвдВЛЖЯИФБфЕФЁЃвђДЫЃЌВЛЭЌжъЯЕжЎМфЕФвХДЋБфвьПЩФмВЛНіНіЪЧгЩЕуЭЛБфКЭDNAжиХХЕМжТЕФЃЌетОЭДгИљБОЩЯИјЭМЮЛПЫТЁЙЄГЬдьГЩСЫбЯжиЕФЮЪЬтЁЃОйР§РДЫЕЃЌШчЙћдкСНИіжъЯЕжЎМфЗЂЩњЕЙзЊЕФвЛИіДѓдМ500 KbЕФађСаБЛгУгкаЮГЩЕФвЛИізїЭМШКЬхЃЌЫљгаЗЂЩњдкетИіЕЙзЊФкЕФжизщЪТМўНЋВњЩњВЛг§ЕФМѕЪ§ЗжСбВњЮяЁЃвђДЫЃЌВЛПЩФмдкетИіЕЙзЊађСаФкЖдЭЛБфНјаазїЭМЁЃЕНФПЧАЮЊжЙЃЌЗЂЩњдкГЃМћжъЯЕжЎМфЕФетбљЕФDNAжиХХЛЙУЛгаБЛБЈЕРЙ§ЃЌШЗЪЕгІИУЪЧетбљЃЌвђЮЊЫќУЧКмФбБЛМьВтЕНЁЃдквЛИізїЭМЪЕбщжаЃЌЫќУЧЕФГіЯжНЋКмгаПЩФмБЛКіЪгжБЕНзюКѓвЛВНЁЃ
гаЪБКђЃЌT-DNAВхШыКЭЗјЩфвВБЛЙлВьЕНФмЕМжТDNAЕФжиХХЃЈShirleyЕШЃЌ1992ЃЛNacryЕШЃЌ1998ЃЛLaufsЕШЃЌ1999ЃЛOgasЕШЃЌ1999ЃЉЁЃвђДЫЃЌЕББЛзїЭМЕФЭЛБфЪЧгЩетаЉЗНЗЈВњЩњЕФЪБКђЃЌРрЫЦЕФРЇФбвВгаПЩФмВњЩњЁЃЕЋдкетаЉЧщПіЯТЃЌжСЩйгавЛЖЈЕФПЩФмадЭЛБфЪЧКЭжиХХЕФвЛИіЛђСНИіЖЯСбЕугаЙиЁЃ
4 ЧАОАеЙЭћ
ФПЧАЃЌдкФтФЯНцжаЕФЭМЮЛПЫТЁвбОВЛНіНіЪЧвЛаЉзЈзЂЕФЃЈКЭГжОУЕФЃЉзЈМвЕФЙЄзїСЫЃЌЖјЪЧУПИіШЫЖМФмЭъГЩЕФЙЄзїЁЃдкЙ§ШЅЕФМИФъжаЃЌВњЩњСЫКмЖрБувЫЕЋЙІФмЧПДѓЕФЙЄОпЃЌЭЌЪБвВгаДѓСПЕФаХЯЂБЛЪеМЏдкУтЗбЕФЪ§ОнПтжаЁЃРћгУетаЉзЪдДЃЌФПЧАДѓВПЗжЕФЭМЮЛПЫТЁЙЄГЬгІИУЪЧПЩвдПЯЖЈЕФЃЌжБНгЕФЃЌвВЪЧМђЕЅЕФЁЃЫцзХЮвУЧЖдФтФЯНцЛљвђзщНсЙЙКЭБфЛЏЕФШЯЪЖЕФдіГЄЃЌЧщПіНЋНјвЛВНИФЩЦЃЌвђЮЊетНЋгажњгкЮвУЧЯћГ§ВПЗжЩЯУцЬсЕНЕФШдШЛДцдкЕФИДдгЧщПіЃЌЛђепжСЩйЪЙЕУЫќУЧПЩБЛПижЦЁЃ
ВЮПМЮФЯз
1. Choe,S., Schmitz,R.J., Fujioka,S., Takatsuto,S., Lee,M.O., Yoshida,S., Feldmann,K.A., and Tax,F.E. (2002). Arabidopsis brassinosteroid-insensitive dwarf12 mutants are semidominant and defective in a glycogen synthase kinase 3 beta-like kinase. Plant Physiology 130, 1506-1515.
2. Gonzalez-Guzman,M., Apostolova,N., Belles,J.M., Barrero,J.M., Piqueras,P., Ponce,M.R., Micol,J.L., Serrano,R., and Rodriguez,P.L. (2002). The short-chain alcohol dehydrogenase ABA2 catalyzes the conversion of xanthoxin to abscisic aldehyde. Plant Cell 14, 1833-1846.
3. Jander,G., Norris,S.R., Rounsley,S.D., Bush,D.F., Levin,I.M., and Last,R.L. (2002b). Arabidopsis map-based cloning in the post-genome era. Plant Physiology 129, 440-450.
4. Lukowitz,W., Gillmor,C.S., and Scheible,W.R. (2000c). Positional cloning in arabidopsis. Why it feels good to have a genome initiative working for you. Plant Physiology 123, 795-805.
5. Michaels,S.D. and Amasino,R.M. (1998). A robust method for detecting single-nucleotide changes as polymorphic markers by PCR. Plant J. 14, 381-385.
6. Nam,H.G., Giraudat,J., Den Boer,B., Moonan,F., Loos,W., Hauge,B.M., and Goodman,H.M. (1989). Restriction Fragment Length Polymorphism Linkage Map of Arabidopsis thaliana. Plant Cell 1, 699-705.
7. Peters,J.L., Cnudde,F., and Gerats,T. (2003). Forward genetics and map-based cloning approaches. Trends in Plant Science 8, 484-491.
8. Rafalski,A. (2002). Applications of single nucleotide polymorphisms in crop genetics. Curr. Opin. Plant Biol. 5, 94-100.