

DNA中的鸟嘌呤容易被活性氧自由基羟化成为82羟基鸟嘌呤,这种碱基改变能诱导GCyTA转化,已证实DNA氧化损伤与心血管疾病!老化等慢性病有关。因此生物体存在预防82羟基鸟嘌呤所致遗传结构完整性损害的防御机制。
8——羟基鸟嘌呤内切酶同时具有82羟基鸟嘌呤糖基酶活性及腺嘌呤核酸内切酶活性,该酶分两步修复DNA中82羟基鸟嘌呤,先将82羟基鸟嘌呤当成自由碱基移去,然后在腺嘌呤位点切开磷酸二酸键的3c和5c端。
最近发现大肠埃希菌的82羟基鸟嘌呤内切酶(糖基酶)能受氧张力增高的诱导,该酶的适应性反应使其在氧化刺激下能发挥更有效的对抗DNA氧化损伤的作用。至今尚未见有哺乳类组织82羟基鸟嘌呤糖基酶诱导的研究。因此,我们尝试在大鼠经KBrO3处理后,其肝和肾中82羟基鸟嘌呤糖基酶的活性改变。KBrO3是一种选择性作用于肾脏的氧化性致癌物。在本研究中,仅发现肾脏82羟基鸟嘌呤糖基酶的活性被氧化诱导,而肝中未被诱导。肾脏中活性增高呈现剂量和时间依赖性。
1 材料和方法
1.1 寡脱氧核苷酸作为底物用于82羟基鸟嘌呤糖基酶的寡脱氧核苷酸的合成按照Kuchino等描述的方法(1987年)。
1.2 以[14C]82羟基鸟嘌呤作为内标物82羟基鸟嘌呤定量的内标物[14C]82羟基鸟嘌呤用[14C]鸟嘌呤制备,按照Degan等描述的程序(1991年)。
1.3 以KBrO3作用于动物0.9%KBrO3盐水通过腹膜内注射雄性Fisher大鼠,大鼠体重约120g。给药前大鼠禁食一夜,为观察KBrO3作用的时间效应,采用80mgPkg剂量;按
24h时间间隔杀死大鼠。为观察KBrO3作用的剂量效应,按不同剂量,最高至160mgPkg给药,给药后9h杀死,分离其肾和肝来制备组织抽提物。
1.4 组织提取物制备
取出大鼠的肾和肝用0.9%冷盐水冲去残血,肾!肝用液氮冷却贮存于-70e备用,用2倍体积的匀浆缓冲液研磨和匀浆冷冻器官,悬浮的匀浆用
15%的链霉素混合,以除去核酸,离心后上清液透析,用作82羟基鸟嘌呤糖基酶分析的组织提取物。上述步骤在4e以下操作。
1.5 酶分析
将4pmol双链DNA底物同5mg上述肾和肝的提取物在30e下孵化2h,1ml反应混合物包括50mmolPLTris2HCl!50mmolPLKCl及1mmolPLEDTA,pH值为7.5,通过90e加热3min来终止反应,按6000rPmin离心10min,取900Ll上清液加100Ll约400次Pmin14C82羟基鸟嘌呤内标液,该混合物加入经82羟基鸟嘌呤单克隆抗体制备的免疫亲和柱。经纯化的
82羟基鸟嘌呤溶于50Ll水,注入5Lm,416mm@25cmHPLC柱,进行高效液相色谱!电化学检测分析,测量电化学检测器上显示的82羟基鸟嘌呤峰高及82羟基鸟嘌呤洗提液的放射活性,峰高用于82羟基鸟嘌呤的定量,是反应介质和加入14C282羟基鸟嘌呤的总和。从总量中减去内标物即得到原反应介质中82羟基鸟嘌呤的量。所加14C282羟基鸟嘌呤可通过校准曲线比较其峰高与放射活性来定量。洗提液的放射活性可用来定量在免疫亲和柱纯化过程中82羟基鸟嘌呤的损失量。加入1ml反应介质中的82羟基鸟嘌呤的量可利用损失率和反应介质中82羟基鸟嘌呤的原始量来测定,酶活性可表达为在上述反应条件下DNA底物中释放出来的82羟基鸟嘌呤的含量,数据可表达为6次实验的均值±标准误。统计分析用t检验。
2 结果
KBrO3对82羟基鸟嘌呤糖基酶活性的影响效应可通过肾与肝中含量的时间及剂量效应来表达。为比较起见,肾脏被选作靶器官,肝脏作控制对比。图1显示腹膜内给80mgPkg剂量KBrO3后,两种器官中82羟基鸟嘌呤含量的时间效应关系,在肾脏,发现其活性在3h[(0.73±0.14)pmol]时显著升高,相比之下,在0h活性只有(0.20±0.06)pmol,而在6h时,其活性达到高大值(125±0.14)pmol,该值是0时的6倍。然后,活性开始下降并在12h回到0h水平;作为对比,肝中活性在相同时间段并无显著变化。图2显示其活性与KBrO3剂量的关系,不加处理时,活性只有(0.20±0.06)pmol,而在20!40!80及
160mgPkgKBrO3腹膜下给予时,其活性分别是(0.33±0.09)!(0.53±0.10)!(0.75±0.13)及(1.30±0.08)pmol,在40mgPkg以上时活性显著增高,且显示剂量反应关系,而肝中活性在相同的剂量范围内并无显著变化。
3 讨论
DNA氧化损伤与心血管疾病!老化等慢性病有关。本研究显示哺乳动物82羟基鸟嘌呤糖基酶也能被氧化作用所诱导,KBrO3作用于大鼠后,肾中该酶活性的增高呈现时间与剂量效应关系(图.,2),而在肝中则不出现该变化。氧化能被SOD酶!过氧化氢酶等所阻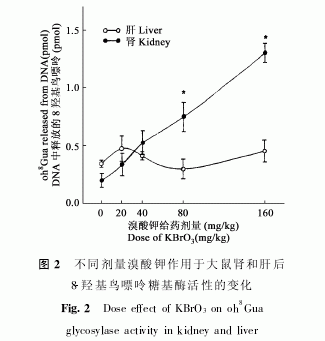
止,也能被氧化修复酶所纠正,如胸腺嘧啶乙二醇糖基酶,82羟基鸟嘌呤修复酶,核酸内切酶等,目前关于氧化修复酶的研究尚少,本研究显示:82羟基鸟嘌呤糖基酶是可诱导的氧化修复酶。
有证据显示需氧生物82羟基鸟嘌呤修复作用的重要性,如DNA中82羟基鸟嘌呤修复酶是普遍存在的,从细菌到哺乳动物的试验中都能发现其作用。
DNA中82羟基鸟嘌呤能诱导GCyTA转化,当在12位密码子上包含82羟基鸟嘌呤的c2Ha2ras基因转染进良性细胞时在该位点诱导GCyTA转化,同时将被转染的良性细胞变成了恶性细胞。
有报道表明,细胞水平的82羟基鸟嘌呤修复活动与致突变作用的诱导与预防有密切关系。由此可见,82羟基鸟嘌呤的修复,对氧化条件下可能造成的遗传结构完整性起保护作用。提示82羟基鸟嘌呤糖基酶的诱导能力是保护DNA免遭氧化损伤的有效生理机制。
总之,哺乳动物细胞中82羟基鸟嘌呤糖基酶能被氧化作用所诱导,结合在大肠埃希菌中已发现该酶的诱导,说明在所有需氧细胞中82羟基鸟嘌呤糖基酶的适应性反应是一种很普遍的现象。普遍发现这种适应性反应表明,82羟基鸟嘌呤的修复对于机体在有氧环境中的生存是一个重要过程。为进一步了解该酶的诱导机制,可研究以下方面问题,如其mRNA水平的改变,弄清诱导启动信号等。